Latitudinal gradients in species diversity in the arctic
Contents
- 1 Introduction Latitudinal gradients suggest that species diversity in the arctic are sensitive to climate. The steep temperature (Latitudinal gradients in species diversity in the arctic) gradient that has such a strong influence on species diversity (Species richness) occurs over much shorter distances in the Arctic than in other biomes.
- 2 Plants
- 3 Animals
- 4 Microorganism
Introduction Latitudinal gradients suggest that species diversity in the arctic are sensitive to climate. The steep temperature (Latitudinal gradients in species diversity in the arctic) gradient that has such a strong influence on species diversity (Species richness) occurs over much shorter distances in the Arctic than in other biomes.
Plants
 Fig. 1. Relationship between mean July temperature and (a) the number of nesting bird species in western and middle Siberia; (b) the number of ground beetle species in local faunas of the Taymir Peninsula; and (c) the number of day butterfly species in the middle Siberian and Beringian areas of the Arctic.
Fig. 1. Relationship between mean July temperature and (a) the number of nesting bird species in western and middle Siberia; (b) the number of ground beetle species in local faunas of the Taymir Peninsula; and (c) the number of day butterfly species in the middle Siberian and Beringian areas of the Arctic. The number of vascular plant species declines five-fold from south to north on the Taymir Peninsula. Summer temperature is the environmental variable that best predicts plant diversity in the Arctic. Other factors are also important, however: regions at different latitudes that have a similar maximum monthly temperature often differ in diversity. Taymir biodiversity values are intermediate between the higher values in Chukotka and Alaska, which have a more complicated relief, geology, and floristic history, and the lower values in the eastern Canadian Arctic with its impoverished flora resulting from relatively recent glaciation. All diversity values on the Yamal Peninsula are even lower than in Canada because of a wide distribution of sandy soils and perhaps its young age. Similar patterns are observed with butterflies (Fig. 1c) and spiders. Therefore, latitudinal gradients of species diversity are best described as several parallel gradients, each of which depends on summer heat, but which may differ from one geographic region to another. This must be taken into consideration when projecting future changes in biodiversity. Figure 1b illustrates how current bioclimatic distributions are related to climate change scenarios by plotting the likely changes in the number of ground beetles for three time slices of mean July temperature derived from the mean of the scenarios generated by the five Arctic Climate Impact Assessment(ACIA)-designated models.
At the level of the local flora (the number of species present in a landscape of about 100 square-kilometers [km2]), there is either a linear or an "S"-shaped relationship between summer temperature and the number of species (Fig. 2). The number of species is least sensitive to temperature near the southern margin of the tundra and most sensitive to temperatures between 3 and 8°C. This suggests that the primary changes in species composition are very likely to occur in the northern part of the tundra zone and in the polar desert, where species are now most restricted in their distribution by summer warmth and growing season length. July temperature, for example, accounts for 95% of the variance in number of vascular plant species in the Canadian Arctic, although extreme winter temperatures are also important. Summer warmth, growing-season length, and winter temperatures all affect the growth, reproduction, and survival of arctic plants. The relative importance of each varies from species to species, site to site, and year to year.
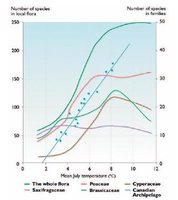 Fig. 2. Relationship between mean July temperature and the number of vascular plant species in local floras of the Taymir Peninsula and the number of vascular species in the local flora of the Canadian Archipelago.
Fig. 2. Relationship between mean July temperature and the number of vascular plant species in local floras of the Taymir Peninsula and the number of vascular species in the local flora of the Canadian Archipelago. North of the treeline in Siberia, mean July temperature decreases from 12 to 2°C over 900 km, whereas mean July temperature decreases by 10°C over 2,000 km in the boreal zone, and decreases by less than 10°C from the southern boreal zone to the equator[1]. Comparing the temperature decrease of 10°C with the projected 2.5°C increase in mean July temperature in the Arctic by 2080 (mean of the two extremes – 1.1 and 4.2°C – projected by the five ACIA-designated models) suggests that much of the Arctic is very likely to remain within the arctic summer climate envelope (although the increase in winter temperature is projected to be higher).
Because of the steep latitudinal temperature gradients in the Arctic, the distance that plants must migrate in response to a change in temperature is much less in the Arctic than in other biomes, particularly where topographic variations in microclimate enable plants to grow far north of their climatic optima. The low solar angle and presence of permafrost make topographic variations in microclimate and associated plant community composition particularly pronounced in the Arctic. Thus, both the sensitivity of arctic species diversity to temperature and the short distance over which this temperature gradient occurs suggest that arctic diversity is very likely to respond strongly and rapidly to high-latitude temperature change.
Latitudinal patterns of diversity differ strikingly among different groups of plants (Table 1). Many polyzonal, boreal, and hypoarctic species have ranges that extend into the Arctic. Some of these (e.g., the moss Hylocomium splendens and the sedges Eriophorum angustifolium and E. vaginatum) are important dominants within the Arctic. Tussocks of E. vaginatum structure the microtopography of broad areas of tussock tundra , and H. splendens exerts a control over nutrient cycling. Tall willow (Salix spp.) and alder (Alnus fruticosa) shrubs as well as dwarf birch (Betula exilis and B. nana) form dense thickets in the southern part of the tundra zone and often have outlier populations that extend far to the north in favorable habitats.
Animals
Latitudinal patterns of diversity in arctic animals are similar to those described for arctic plants. Species diversity declines in parallel with decreasing temperature in most animal groups (Fig. 1), including birds, ground beetles, and butterflies. However, in some groups (e.g., peatland birds and sawflies at local sites in the European Arctic), concentrations of both species diversity (Species richness) and density per unit area can increase compared with more southern territories, perhaps because the habitat types appropriate to these groups are more diverse in the tundra than in the boreal forest. In general, the latitudinal decline in the number of animal species is more pronounced (frequently greater than 2.5-fold) than in vascular plants. As with plants, at a given temperature there are more animal species in Beringia, with its complicated relief, geology, and biogeographic history than in the Taymir Peninsula. Many animal species are restricted to the boreal zone because they depend on the crown, wood, roots, or litter of trees, which are absent in the tundra zone. These groups include wood-boring insects and wood-decaying fungi and their predators, as well as mammals and birds that specialize on tree seeds and leaves. Other important animals, including the raven (Corvus corax), wolf, red fox (Vulpes vulpes), and ermine (Mustela erminea), are primarily boreal in distribution but remain an important component of many arctic ecosystems. There are a few terrestrial animals restricted to the high Arctic such as the sanderling (Calidris alba) and a common Collembola, Vertagopus brevicaudus. Other arctic species have their centers of distribution in the northern, mid- or southern Arctic (Table 1). The more diverse patterns of animal distribution compared to plants make it more difficult to project how animals will respond to climatic warming. Some herbivores have distributions that are more limited than those of their host plants, so it is possible that warming will allow these species to extend northward relatively rapidly.
As with plants, latitudinal patterns of diversity differ strikingly among different groups of animals (Table 1). The common species tend to be more broadly distributed in the far north. In northern Taymir, there are only 12 species of springtails but 80% of these occur in all microsites and topographic locations investigated. Some boreal birds, such as the American robin (Turdus migratorius), penetrate only into the southern part of the tundra while others can occur far from the area of their continuous distribution (climatic optimum): in the vicinity of Dixon (Taymir), the forest thrushes T. pilaris and T. iliacus form populations in the northernmost part of the tundra zone, 400 km from the last outposts of the forests. At the southern limits of the tundra, there is greater specialization among microhabitats. Many more species occur in intrazonal habitats, occupying relatively small and isolated sites, than in zonal habitats that contain only a small proportion of the regional fauna. Warming is therefore likely to lead to more pronounced habitat and niche specialization.
An important consequence of the decline in numbers of species with increasing latitude is an increase in dominance. For example, one species of collembolan, Folsomia regularis, may constitute 60% of the total collembolan density in the polar desert[2]. These "super-dominant" species are generally highly plastic, occupy a wide range of habitats, and generally have large effects on ecosystem processes. Lemmings (Lemmus spp. and Dicrostonyx spp.) are superdominant species during peak years of their population cycles[3] and have large effects on ecosystem processes[4].
Microorganism
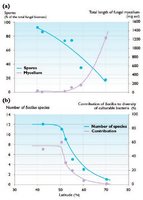 Fig. 3. Latitudinal distribution of (a) soil fungi and (b) bacilli; curves are hand-fitted (recalculated from data in Mirchink.
Fig. 3. Latitudinal distribution of (a) soil fungi and (b) bacilli; curves are hand-fitted (recalculated from data in Mirchink. Arctic soils contain large reserves (standing crops) of microbial (mainly fungal) biomass, although the rate of microbial growth is generally lower than in the boreal zone. Surprisingly, under severe arctic conditions, soil microbes fail to produce spores and other dormant structures that are adaptations to harsh environments (Fig. 3). The species diversity of all groups of soil microorganisms is lower in the Arctic than further south, decreasing from about 90 species in Irish grassland, through about 50 species in Alaskan birch forest, to about 30 species in Alaskan tundra. As with plants and animals, there are large reductions in numbers of microbial species with increasing latitude, although these patterns are less well documented. A correlate of the decreasing number of species with increasing latitude is increasing dominance of the species that occur, as with plants and animals. One yeast (Cryptococcus laurentii), for example, constitutes a large proportion of yeast biomass across a range of community types in the northern Taymir Peninsula.
The hyphal length of fungi in the Arctic shows a latitudinal trend in which the abundance of fungi, as measured by hyphal length, decreases toward the north. Although it is not known if this trend also applies to the species diversity of fungal mycelia (the belowground network of fungal filaments or hyphae), it is clear that the amount of fungal hyphae is low in the Arctic. In the high Arctic, fungal hyphal length was 23±1 m/g in a polar semi desert on Svalbard (78°56' N), 39 m/g on a beach ridge, and 2228 m/g in a mesic meadow on Devon Island (75°33' N). At Barrow, Alaska, hyphal length was 200 m/g. In a subarctic mire in Swedish Lapland, hyphal length was 3033 m/g. These values can be compared with 6,050 to 9,000 m/g in temperate uplands in the United Kingdom and 1,900 to 4,432 m/g in temperate woodland soils.