Evolution
Contents
- 1 Introduction Evolution is most commonly defined as a change in allelic (varieties of genes) frequencies in a population (Evolution) over time. In other words, evolution encompasses a series of mechanisms that lead to changes in the relative proportion of different types of genes contained in a population where these changes persist from one generation to another. The primary mechanisms that result in such changes are selection, genetic drift, mutation, and gene flow.
- 2 Mechanisms of evolution
- 3 Limitations on evolution
- 4 Putting it all together
- 5 Evolution at multiple levels: micro to macroevolution
- 6 Further reading
Introduction Evolution is most commonly defined as a change in allelic (varieties of genes) frequencies in a population (Evolution) over time. In other words, evolution encompasses a series of mechanisms that lead to changes in the relative proportion of different types of genes contained in a population where these changes persist from one generation to another. The primary mechanisms that result in such changes are selection, genetic drift, mutation, and gene flow.
It is a common misconception that evolution occurs only through selection, but all four mechanisms often co-occur and selection is not necessarily the dominant force of evolutionary change. As Charles Darwin is recognized as the preeminent scientist to write formally about evolution by selection, this mechanism is often termed Darwinian selection. In fact, many scientists were synthesizing their ideas about evolution at the same time as Darwin and the first formal presentation of evolution by natural selection was jointly authored by Alfred Russell Wallace and Charles Darwin. However, Darwin’s seminal book entitled On The Origin of Species by Means of Natural Selection is often cited as the genesis of evolutionary theory as it contains a litany of evidence indicating how selection operates and how species are descended from common ancestors.
Darwin’s theories on evolution lacked any knowledge of genetic change and it was not until Gregor Mendel’s work on the genetic heritability of pea plant characteristics were incorporated into evolutionary thinking that the new synthesis of evolutionary theory was born. This new synthesis led to a flurry of research in the 1920s to 1960s that described and formalized the genetic mechanisms of evolution. Evolutionary theory has changed substantially since the initial days of Darwin and Wallace.
As there are now tens of thousands of scientific studies that demonstrate how evolution works, its characterization as a “theory” is somewhat misleading, and many scientists feel the term theory misrepresents the overwhelming weight of evidence that supports the modern conception of evolution and how it works.
Mechanisms of evolution
Selection
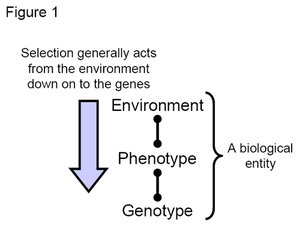 Figure 1. Cartoon indicating that selection acts from the environment through the phenotype to affect the genotype. (Credit: John Swaddle)
Figure 1. Cartoon indicating that selection acts from the environment through the phenotype to affect the genotype. (Credit: John Swaddle) If particular varieties of genes (alleles) confer a fitness advantage then these genetic varieties will increase in frequency from one generation to another via selection. A fitness advantage occurs when an individual produces relatively more descendants (i.e. offspring) that survive to reproductive age than other individuals in the population. Hence, genes of individuals producing the most offspring will be over-represented in the next generation, leading to a change in overall gene frequencies in favor of those selected alleles. Key to this process is the presence of heritable (i.e. at least partially genetically determined) variation in the population.
Selection generally acts from the environment through the phenotype (the physical manifestation of the underlying genes) to the genotype (the genetic make-up of the individual) (Figure 1). Hence, understanding how phenotype relates to genotype (i.e. understanding developmental and genetic processes) is key to understanding how selection operates in any population.
Selection can be visualized by the relationship between values of a genetically heritable trait (a characteristic of individuals in the population, for example, body size) and a metric of fitness (Figure 2). The genes that code for the higher fitness trait values will increase in frequency in the population from one generation to another. Although it is tempting to view the relationship between the heritable trait and fitness as linear (Figure 3a), in reality this relationship is often nonlinear and commonly shows a humped distribution (Figure 3b).
 Figure 2. Graph indicating relationships between variation in a heritable trait with a measure of fitness. The red line indicates a trait under positive directional selection. The green line represents a trait which shows neutral variation. (Credit: John Swaddle)
Figure 2. Graph indicating relationships between variation in a heritable trait with a measure of fitness. The red line indicates a trait under positive directional selection. The green line represents a trait which shows neutral variation. (Credit: John Swaddle) If fitness increases in proportion to the size of the heritable trait, the trait is described as experiencing directional selection. In Figure 3a, this trait is under positive directional selection. If the slope of the line was negative (i.e. smaller trait values conferred a fitness advantage) then the trait would be under negative directional selection. With these examples, the steepness of the line is a measure of the strength of selection, with steeper slopes indicating stronger selection. If the slope is zero (i.e. a flat line) then there is no selection and the phenotypic trait is said to display neutral variation (Figure 2).
As described above, under directional selection the mean value of the heritable trait will change over generations, shifting in either a positive or negative direction to match the direction of the selection pressure (lower panels of Figure 3a). This is not the case under stabilizing selection, where the average trait value confers a fitness advantage (Figure 3b). In this situation, the population will converge on values closer to the population mean over subsequent generations. Hence, the population mean value for the heritable trait will not change over time but the variance (a measure of the relative spread of trait values around the population mean) will decrease (lower panels of Figure 3b). This result is important for two reasons. First, it demonstrates that evolution does not necessarily lead to changes in average traits and characteristics over time—traits do not need to get smaller or larger over time to be under selection. Second, stabilizing selection explicitly erodes genetic variation in the population, as the variance in the heritable trait gets smaller and smaller over time. As genetic variance is a necessity for selection to operate (imagine altering Figures 2 and 3 so that all individuals have the same trait value—there would be no capacity for selection to work), stabilizing selection can greatly influence the capacity for future evolutionary change.
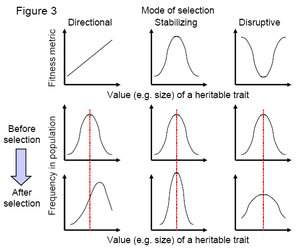 Figure 3. Each column of graphs indicates the consequences of the three major forms of selection: directional, stabilizing, and disruptive selection. For each column, the upper graph indicates how fitness varies with trait value; the middle graph indicates the frequency of trait values in the population before selection acts (note that all the populations start from the same point); and the lower graph represents how the trait values change after one generation of selection. (Credit: John Swaddle)
Figure 3. Each column of graphs indicates the consequences of the three major forms of selection: directional, stabilizing, and disruptive selection. For each column, the upper graph indicates how fitness varies with trait value; the middle graph indicates the frequency of trait values in the population before selection acts (note that all the populations start from the same point); and the lower graph represents how the trait values change after one generation of selection. (Credit: John Swaddle) Stabilizing selection is a common occurrence in nature. Disruptive selection (Figure 3c) is more rare. Under disruptive selection, the two extreme limits of trait values confer fitness advantages. Hence, the population will tend to split over time, with two groups of individuals emerging—one with the lowest trait values and one with the highest trait values. If disruptive selection persists for sufficient time, there will be very few individuals with the average trait value and the population frequency distribution will become bimodal, resulting in two sub-populations. Therefore, disruptive selection is important to speciation processes as it can separate subgroups within a population.
Sexual selection
Many evolutionary biologists would include assortative mating (i.e. nonrandom mating, often indicating that males and females are choosing and/or actively competing for mates) as a mechanism of evolution, but this process affects evolution only when the assortative mating leads to fitness differences. That is when there is selection for differential mating success—a subset of natural selection termed sexual selection. Sexual selection often operates by one sex (e.g. the female) selectively choosing to mate with a particular kind of male—therefore, the fitness of those types of males increases. Or males (or females) can compete with each other to gain access to the opposite sex. Either way, there may be particular characteristics of males and females that make them more attractive as partners or more competitive at gaining access to mates. Such traits are said to be under sexual selection. Famous examples of sexually selected traits are the elaborate tails of peacocks or the huge antlers of deer. Such traits may substantially increase mating success but they can also lower some other aspects of fitness, such as the ability to get away from predators. Therefore, sexual selection is often viewed as being in conflict with other components of natural selection.
Genetic drift
Drift refers to a suite of random (or stochastic) processes involved in producing the next generation of individuals. Such processes can be at the genetic level, for example, the (mostly) random chance of which copies of alleles make it into the germ (sperm and egg) cells through meiotic reduction division. In many sexually reproducing organisms, cells contain two copies of alleles (diploid) at every locus because they possess paired chromosomes. But because the next generation is formed by the fusion of a sperm from the father and an egg from the mother, germ cells go through a random halving of these chromosome pairs (i.e. just one copy of each chromosome makes it into the sperm or egg) so they can fuse to form a zygote (viable next generation cell) that has the same number of chromosomes as the adults. During this meiotic reduction division, it is almost random which copy of the alleles makes it into the sperm and eggs. This means that sperm and eggs within the same individual are rarely genetically identical, which is why siblings are different to each other even though they have the same parents. The meiotic shuffling of genes changes gene frequencies from one generation to another at random—some alleles are lucky enough to make it into the sperm that fertilizes the egg, whereas other alleles are unlucky and are lost.
A useful analogy to this genetic drift process is tossing a coin. Every toss of the coin determines which of two alleles at one locus makes it into the next generation. Imagine starting a population of ten individuals, which would mean tossing the coin ten separate times. The first time you do this, you may get 6 heads and 4 tails (or something different). In allele frequency terms, this means an allelic frequency of heads as 0.6 (6 out of 10). The second time you do this from the same coin (i.e. exactly the same starting point), you will likely get some other ratio of heads to tails. For example, 3 heads and 7 tails, which translates to an allelic frequency of heads as 0.3 (3 out of 10). As you can see, allelic frequencies can change substantially by random chance.
Figure 4 shows some typical changes in allelic frequencies under genetic drift (for one locus with two alleles) where generations are tracked on the horizontal axis and allele frequency (ranging from 0 to 1) on the vertical axis. If the allele frequency reaches zero it is lost forever. For each graph, five populations have been simulated, with each independent population indicated by different color lines. Drift is typified by a random walk in allele frequencies over generations. At small population sizes (akin to tossing a coin a small number of times) there are larger swings in allele frequency because it is not that improbable to toss all heads (i.e. obtain an allele frequency of 1) if you toss a coin three times only (i.e., a population size of 3). An example of drift in a small population (with 10 individuals) is depicted in Figure 4a. Note that there are several incidences were one of the alleles is lost due to chance events. However, at large population sizes drift has less effect and the swings in allele frequency are much smaller. To keep with the analogy, if you toss the coin a thousand times the chances of tossing all heads are very remote. Hence, genetic drift can be a very influential evolutionary force at small population sizes. An example of drift in a large population is described in Figure 4b. Note that in there is a much lower probability of an allele being lost under drift at a large population size.
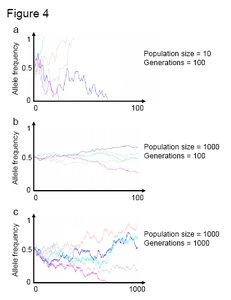 Figure 4. Graphs of allele frequency over time for a one-locus two-allele system experiencing genetic drift. Each line on a graph represents a different population. As drift is a random process, each population shows a different path over time even within the same graph. If a line reaches zero or 1 it cannot "bounce" back as the allele is either lost from the population or becomes fixed in the population (resulting in the permanent loss of the complementary allele). Note that genetic drift has larger effects on allele frequency at smaller population sizes and over longer time periods. (Credit: John Swaddle)
Figure 4. Graphs of allele frequency over time for a one-locus two-allele system experiencing genetic drift. Each line on a graph represents a different population. As drift is a random process, each population shows a different path over time even within the same graph. If a line reaches zero or 1 it cannot "bounce" back as the allele is either lost from the population or becomes fixed in the population (resulting in the permanent loss of the complementary allele). Note that genetic drift has larger effects on allele frequency at smaller population sizes and over longer time periods. (Credit: John Swaddle) Drift can also have a large influence on allele frequency over long time periods. In the context of our coin tossing analogy, if you keep tossing a coin over and over again, eventually you will get a long run of heads that would substantially change allele frequencies. Example patterns of drift over extended time periods are given in Figure 4c. Note that alleles can be lost by chance over long time periods even if the population is large.
Overall, small, fragmented populations are sensitive to drift. Likewise, evolution over long time periods (e.g. as studied in the fossil record) will also be commonly be influenced by drift processes.
Drift is not limited to genetic processes—which individuals get to breed to form the next generation of a population can also be random. For example, imagine a population on an island where there is a fierce fire, which wipes out 90% of the individuals. Which individuals survive may be randomly determined, but if these surviving 10% are not entirely representative of the starting 100%, allele frequencies will change in the next generation because of the random bottleneck that the population just experienced. Such population bottlenecks are very important to consider for endangered species (IUCN Red List Criteria for Endangered) as the bottleneck often dramatically reduces genetic variance in the species, which limits how the species can evolve and respond to further environmental change. For example, cheetahs have experienced such a severe bottleneck that scientists estimate that all surviving cheetahs across the globe are more closely genetically related to each other than siblings usually would be in a normal population. As you can imagine, this creates terrible problems with inbreeding and rare genetic disorders. These problems have been largely brought about by the random evolutionary process of drift.
A similar individual-level drift process occurs when new founding [[population]s] are formed—this is called the “founder effect”. The prospecting individuals that form a new population isolated from the original group can be randomly different to the larger group. This has been observed time and time again in ecology and famously documented in human populations. For example, founding Amish populations in North America did not carry alleles at the same frequency as the population they left behind in Europe and, by chance, carried some rare genetic disorders which are still prevalent at dramatically higher frequencies in Pennsylvania than in other parts of the world. Because a small number of individuals founded these populations the evolutionary force of drift has created substantial health problems in modern day humans.
Mutation
Genetic variation is the fuel for evolution. Without genetic variation there will be no change in allelic frequencies as there is nothing to change to. One of the principle ways in which genetic variation is introduced and maintained in populations is through mutation. This is a catch-all term that refers to heritable changes in genetic information, where these changes can happen at the small scale of single nucleotides (i.e. acid base pair components of DNA) up to large scale restructuring, loss, and gain of whole chromosomes (organized strings of genetic information).
Mutation generally happens as a mistake during regular cell copying and repair mechanisms, and although the word mutation has negative connotations in everyday society, everyone carries millions of mutations and is the product of previous mutations. If mutations are very severe, often that cell does not function properly. But if mutations are minor (or neutral) mutations can easily persist.
From an evolutionary perspective, the most important mutations are those that occur in the cells that produce the gametes (i.e. sperm and egg cells). As it is these gametic cells that pass genetic information to the next generation—hence affecting a change in genetic information if a mutation is passed from parent to offspring. Therefore, a mutation in a gamete cell that forms a viable zygote brings about an evolutionary change, as allelic frequencies will have changed from the parental generation.
Many mutations are neutral (i.e. do not influence the fitness of the individual bearing that mutation), but some will bring about a dramatic change in fitness. Often this will be a decline in fitness but, on occasion, the mutation could bring about an increase in fitness. It is important to remember that the functional consequence of the mutation is not an indication of evolution itself—evolution merely requires a heritable change in the genes, which is brought about by either neutral or functional mutations in viable gametic cells.
Gene flow
Often the most overlooked, but in some ways the most simple, mechanism of evolution is gene flow. This is a general term that refers to the movement of genes from one population to another. In most practical cases, this equates to the movement of individuals from one population to another with the migrants mating with individuals at their new locality. The introduction of new allelic combinations into the offspring from such matings brings about a genetic change.
Evolutionary biologists often assume that a high degree of gene flow among populations will tend to be a homogenizing force. In other words, if genes and individuals are constantly mixing across population borders, then populations will tend to get more and more genetically similar over time. As an analogy, imagine separating a deck of playing cards into the respective suits and keeping those “populations” of hearts, clubs, diamonds, and spades separate to each other. Now allow some “gene flow”, which would mean swapping cards among the piles. This changes the ratio of suits to each other (i.e. evolution is occurring). Now if you picked up all the cards, thoroughly shuffled them, and then put them back in four piles, you have simulated a maximum amount of gene flow, and chances are that you have a pretty even mix of each suit in each of the piles. Hence, increased gene flow has homogenized the populations.
The previous analogy worked because individuals were moving (genes were flowing) at random from one population to another. However, if gene flow is directed or nonrandom, this mechanism can be a force that results in increasing differences between populations. An example of this comes from great tits (small European birds) in Whytham Woods near Oxford University. Here, birds leave their natal sites and settle in different parts of the woods according to their body size—bigger birds move to one part, and smaller birds move to another. Hence, in this example, nonrandom gene flow is bringing about a form of directional evolution.
Limitations on evolution
Although this is not intended to be an exhaustive list of the various factors that limit how evolution works, we have highlighted two broad categories that commonly affect how a population can evolve.
Constraints
All of the forces of evolution work within constraints. We do not intend to list all these possible constraints in detail here but, rather, point to some of the major constraints that any student of evolution should keep in mind as they interpret evolutionary information.
The inherent genetic variation of the parental generation constrains the possible allelic combinations that can be observed in the offspring (plus additional variation from mutation). Hence, genetic variability is a large constraint on how evolution proceeds. No matter how strong a selection pressure there may be you will not be able to select for horses to grow wings, as modern horses simply lack the genetic variance for wing growth. Evolutionary biologists also often refer to phylogenetic constraints—the evolutionary history of common ancestors, reflected in the DNA, limit the ways in which the genome can change in the future.
Another common constraint is the general pattern of developmental processes and stages that individuals have to go through. For example, frogs are constrained to go through a tadpole stage and cannot skip ahead. Only a change to developmental regulator genes could alter such a developmental sequence.
Time is also a considerable constraint on how evolution progresses. As evolution is defined as heritable allelic changes over time, if sufficient time does not pass, evolution may not happen. Evolutionary time is often measured in terms of generations, so “time” in bacterial evolution will pass at a much faster rate than “time” in terms of elephant evolution.
Physical properties also constrain evolution. For example, the much feared giant insects of B movie classics cannot evolve because of the ways in which insects respire and the physical relations between surface area:body size ratios. Insects get oxygen for the bodies through small holes in the surface of their cuticles--these holes are called spiracles. The size of spiracles increases with the surface area of the insect and, hence, increases as a two-dimensional plane of length and breadth. However, insect body size increases in three-dimensions (length, breadth, and height); hence, body size increases more quickly than spiracle area as insects get larger and larger. In other words, larger insects have a smaller surface area relative to their body size than smaller insects. The physical properties of how insects respire mean that insects have an upper limit as to how large they can get, as if they get too large they suffocate as they cannot get enough oxygen through their spiracles to supply their increasing body mass. In the natural world, physical constraints commonly limit how evolution can proceed.
Non-independence of traits
Genes, and their allelic varieties, are commonly not independent of each other. For example, genes physically located next to each other on a chromosome are physically linked and will often be inherited together from parent to offspring. If one of these genes if highly beneficial whereas the other is mildly deleterious (harmful), the deleterious gene can "hitchhike" with its buddy and increase in frequency even though it will lead to the increase in frequency of a maladaptive trait.
A similar phenomenon can occur at the phenotypic level through a genetic property known as pleiotropy. Pleiotropy is when a single gene (or gene complex) influences the expression of multiple phenotypic traits. Pleiotropy is fairly common and results in phenotypic traits being correlated with each other through common genetic mechanisms. Hence, if one traits is highly adaptive but another pleiotopically linked trait is mildly deleterious, the deleterious trait can be selected for because of its pleiotropic association with the more advantageous trait. Again, maladaptive states can evolve because of this form of correlation among traits.
Traits can also be linked to each other through trade-offs in allocation of energy and resources. For example, a growing organism has a set energy budget based on what it eats, and that energy can be allocated to various traits and behaviors, which will trade-off against each other because of limited resources. An organism could invest more in reproductive activities, or more in growth, but cannot maximize both. Hence, the evolution of life history traits (which are often closely associated with fitness) is limited because of trade-offs.
Putting it all together
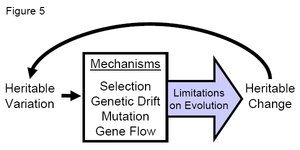 Figure 5. Cartoon indicating the general process of how evolution works. Note the importance of heritable genetic variation—the fuel for evolution. (Credit: John Swaddle)
Figure 5. Cartoon indicating the general process of how evolution works. Note the importance of heritable genetic variation—the fuel for evolution. (Credit: John Swaddle) Although this diagram is a simplification of a very complex and dynamic process, Figure 5 summarizes the major processes involved in the evolution of populations. An evolutionary force operates on the genetic variation in the population, bringing about a heritable change in the next generation, while the evolutionary force works within limitations. As evolution is iterative, this process constantly loops back on itself, bringing about further change in future generations depending on the extent of the evolutionary forces.
Evolution at multiple levels: micro to macroevolution
Most of the descriptions so far have concentrated on evolution at the individual level, with a population defined as a group of individuals. However, for evolution to work we do not need to restrict ourselves to individual-level thinking. As long as the characteristic we are interested in (from an evolutionary perspective) is heritable (i.e. is passed from one generation to another) and is variable (i.e. there are different representations of this trait) then evolution could act. Hence, evolution could happen at the gene-level, as a chromosome could be seen as a population of genes. Such gene-level (reductionist) thinking was popularized by Richard Dawkins in the 1970s and persists in most molecular evolutionary research.
Similarly, a group of individuals could be seen as a single entity (imagine a culture of bacteria growing on a slice of bread) and that group may have a property that cannot be better described at a lower (e.g. individual) level. Population density is a group trait, as it is the property of multiple individuals. So, as long as density is heritable in some fashion—meaning that high density groups tend to give rise to other groups of high density—then this group trait can also evolve. If density was related to a group measure of fitness (i.e. the rate at which a group gives rise to another group) then density may be under group selection. Group selection is often incorrectly defined (especially in the social sciences) as individuals acting for the good of the group—this is not an accurate evolutionary definition of group selection.
As stated a couple of times already, variation is key for evolution to work, and as we climb the organizational ladder of biology from genes to groups and species, we tend to lose variation among entities. For example, if you look at the variation in height among people within a group (perhaps the class you are taking or your sports team) you will find some variation. But if you take the mean height of your group and compare it to the mean height of other groups (perhaps other classes or other teams) you will find much less variation. So, if variation tends to get smaller as you go up the scale of biological organization, evolution tends to be slower and evolutionary forces tend to be weaker at those higher levels. This will usually mean that individual- and gene-level processes dominate over shorter time periods, but it also implies that evolution over longer time periods (e.g. evolution in the fossil record) should be relatively more influenced by group- and species-level evolutionary processes.
The general study of evolution at species levels (and higher) is termed macroevolution and often requires long (generational) time periods to notice any change. The fundamental mechanisms that we have discussed at the population level and below (i.e. microevolution) still apply at macroevolutionary levels but may play out in different ways because of the differences in how macroevolutionary traits are inherited, produced, and defined.
Further reading
- Understanding Evolution website from University of California at Berkeley.
- PBS evolution television series and associated online resources and learning tools, suitable for students and teachers.
- Darwin, C. R. 1859. On the origin of species by means of natural selection, or the preservation of favoured races in the struggle for life. 1st edition. London: John Murray.
- Freeman, S. and Herron, J. C. 2007. Evolutionary analysis. 4th edition. Upper Saddle River, NJ: Pearson Prentice Hall. ISBN: 0132275848.
- Barton, N. H., Briggs, D. E. G., Eisen, J. A., Goldstein, D. B. and Patel, N. H. 2007. Evolution. 1st edition. New York: Cold Spring Harbor Laboratory Press. ISBN: 0879696842.
- Mayr, E. 2001. What Evolution Is. New York: Basic Books. ISBN: 0465044263.
- Coyne, J. A. and Orr, H. A. 2004. Speciation. Sunderland, MA: Sinauer Associates. ISBN: 0878930892.
- Lewin, R. and Foley, R. A. 2004. Principles of human evolution. 2nd edition. Oxford: Blackwell Publishing. ISBN: 0632047046.