Template:Centriole
Contents
Centriole Inheritance Model in Fertilization
Edited: April 26th, 2018
Author: Christopher Fox-Strauss
Topic Editor: Tomer Avidor-Reiss
Topics: Spermatogenesis, centriole, fertilization, embryogenesis, tubulin, mitosis, meiosis
Centrioles are an organelle that serves many functions within cellular processes. The centriole is a microtubule-organizing center that recruits pericentriolar material (PCM) to build a spindle matrix that projects into the cytoplasm. Centrioles appear in couples in each cell in the form of a mother and daughter centriole (Wilson, 2008). Each new cell must have two centrioles, and as a result the centriole must duplicate during each cell cycle in order to fulfill this requirement.
The focus of this article is to provide an overview of the role of centrioles as they relate to fertilization, and more specifically how centrioles in eukaryotic sperm look and function as well as how they duplicate to form centrosomes during embryogenesis. Two competing theories attempt to explain the form and function that sperm's centrioles can be found in: reduction theory and remodeling theory. This article will explain the structures, proteins, and processes involved in spermatogenesis, centriole duplication, and fertilization to help illuminate the concerns with the current reduction theory.
Definitions and Background Information
Definitions of Structures and Definitions of Centrioles
- Centriole polarization- Centrioles are polarized, having both a base and a tip. The base of the centriole contains a cartwheel and the tip associates with the appendages.
- Mother centriole- The mother centriole is the fully matured centriole and possess appendages used to form cilia. The mother centriole will help generate a procentriole during S phase.
- Daughter centriole- The daughter centriole is the juvenile centriole in a cell. It lacks appendages, and is linked to the mother cell by a protein linker chain. The daughter centriole also forms a procentriole during S phase.
- Appendages- Matured centrioles (mother centrioles) posses an additional structure, called appendages. The appendages sprout from the barreled tubulin matrix of the centriole, and just out as distal and sub distal arms. These appendages attach the centriole to the plasma membrane of the cell, allowing the mother centriole to push out a tubulin axoneme that will form the cilia of the cell.
- Protein Linkage- The mother and daughter centrioles are linked by a protein chain to ensure they are together when they start to form a centrosome.
- Cilia- Cilia are hair-like projections from the plasma membrane of the cell that serve countless functions in cells. Often used for transport, many somatic stem cells use their cilia for sensory function.
- PCM (Pericentriolar Material)- PCM is the collection of proteins found around the centriole and is associated with the 9 fold symmetry of the centriole.
- Centrosome: The centrosome is an organelle within the cell assigned the function of forming a astral microtubule matrix required for cell division and fertilization. It is formed from two centrioles, and is surrounded by PCM.
- Microtubule: A microtubule is a very small tubular structure formed from tubulin.
- Astral Microtubule: Astral microtubules are microtubules formed during cell division and fertilization, and originate from the centrosome. They form star-like shaped projections from the centrosome into the lumen and onto the plasma membrane. Astral microtubules are important in organizing the distribution of organelles and cellular volume during divisions.
- Spindle: Spindles are a form of tubulin that extend into the cytoplasm of a cell during mitotic and meiotic division to pull the DNA chromosomes apart into their respective cells.
- Procentriole: A procentriole is the budding new centriole consisting of a cartwheel and microtubules. The procentrioles will work with their parent centriole to form a centrosome during Mitosis. (S-Phase)
- Cartwheel: The cartwheel of a centriole is located on the base of developing centrioles near the parent centriole, and contains 9 spokes. This structure associates with SAS-6 to build the nine-fold symmetry present in the pro-centriole. [1]
- Proximal: This word is used to refer to the region of an object or animal that is on the closer end of the observed object.
- Distal: This word is used to refer to the region of an object or animal that is further projecting or situated from the observed object.
Proteins Involved in Centriole and Centrosome Formation
- Tubulin: is the primary protein present in microtubules, and is thus found in microtubule asters. [2]
- POC5: is required for construction of the distal portion (tip) of a centriole, and for centriole elongation into the lumen. [3]
- SAS-6: has nine dimers associate with the cartwheel of a pronucleus to help form the nine-fold symmetry present in centrioles. [1]
- Centrin: like POC5, is a protein found in the lumen at the tip of centrioles, and thus is found alongside active centrioles. [4]
- POC1B: is important for centriole stability. [5]
- PLK4: is the master regulator of centriole duplication as it stimulates the formation of procenrioles on the surface of parental centrioles. It recruits proteins like SAS-6 and POC1B to form the cartwheel structure at the base of the pronucleus. [6]
Stem Cell Centriole Cycle
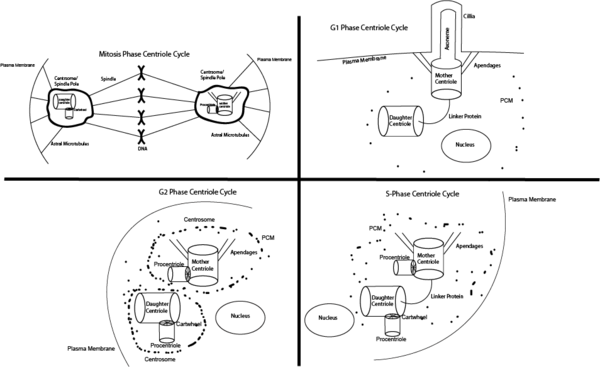
Growth 1 Phase (G1)
During the first stage of cellular growth (G1), the cell possesses two centrioles (a mother and a daughter) bound by a protein linkage. The mother centriole is fully matured, and has appendages that allow the centriole to dock on the plasma membrane. From the mother centriole two of the three microtubules extend from each of its 9 triplets out toward the membrane. This new microtubule structure made of nine doublet microtubules is referred to as the axoneme, and it provides the structure of the projection of the cell. This membrane extension is referred to as the cilia membrane, and is used in numerous ways by different cells types, though often involves sensory functions such as sight or smell.
Synthesis Phase (S)
In Synthesis (S), all DNA and centrioles synthesize a new copy of themselves, and thus duplicate. The importance of this step is to ensure both the mother and the daughter cells have a full set of functional DNA and a mother and daughter centriole. The cilium begins to fall apart, releasing the mother's appendages from the plasma membrane, and while still bound by a linker protein, the centrioles begin to form a procentriole. The procentriole forms near the existing centriole, and helps to form the centrosome.
Growth 2 Phase (G2)
During Growth stage 2 (G2), the linker between the mother and daughter centriole deteriorates, and PCM begins to heavily condense around the centrosomes. This process creates two discrete centrosomes between the centriole pairs. Additionally, the centrosomes begin to project astral microtubules out.
Mitosis (M)
The goal of Mitosis (M) is to have two fully functional cells split from the single one from the previous steps. To achieve this, the centrioles help direct the equal division of DNA by use of spindles coming from the centrosomes. Each centrosome is anchored to the plasma membrane via the astral microtubule, and will work to distribute the genetic material and organelles. Centrosomes help the daughter cells to divide evenly by ensuring each cell has 2 centrioles and the proper organelles. This is accomplished by the astral microtubules which reach out from the centrosome into the cellular lumen and onto the plasma membrane to organize the structure and composition of the daughter cells.
Spermatogenesis
Spermatogenesis is the process of turning the stem cell precursor into a fully functional sperm cell. This process involves the creation of haploid (containing only one set of chromosomes) spermatids. The stem cell undergoes an asymmetrical mitotic division before performing several symmetrical mitotic divisions and then on to meiosis. In the final round of development, the spermatid produces an axoneme just as a stem cell does in Growth 1. The axoneme will later become the sperm's flagella below the sperm's neck. After flagella formation in the matured sperm, and before fertilization of the ovum, the sperm looses a typical centriole. This process is not fully understood, and has led to two competing hypotheses as to how the sperm disassembles the distal centriole.
Two Theories on Centriole Inheritance: Reduction and Remodeling
Definitions of Structures and Names of Fertilization
- Haploid refers to cells (and in particular nuclei) that contain only one set of chromosomes. Two haploid cells will combine during fertilization to form a diploid cells that can undergo cell division and growth.
- Sperm is the haploid male gamete. The sperm contains a single typical centriole at the base of the DNA-containing head. The sperm's long axoneme tail allows for locomotion to find the egg.
- Ovum (also referred to as egg) is the haploid female gamete that contains a pronucleus, releases polar bodies during fertilization, and contains no centrioles. The egg contains the genetic material of the mother, including in eukaryotes, the mitochondrial DNA.
- Polar body are cells that form during meiosis and are excluded from the egg.
- Asymmetric mitosis is the process that occurs when a cell gives rise to two cells of different fates. This occurs with stem cells when the parental stem cell produces a single determined cell and a copy of itself.
- Symmetric mitosis is the process that occurs when the parental cell forms two daughter cells with identical fates. This occurs during normal cell growth and proliferation. Additionally, this process allows for cells to multiply, such as in spermatogenesis.
- Meiosis- Meiosis is the cellular process of forming two haploid gametes from a diploid cell. These gametes are in the form of eggs and sperm in females and males, respectively.
- Zygote- A zygote is a diploid cell formed from the fusion of a haploid egg and haploid sperm. This is what will undergo cleavage in order begin embryo development.
Both hypotheses recognize that cells (eg zygote) that divide require two centrioles, eggs contain no centrioles, and sperm contain only one typical centriole. The next step in embryogenesis is fertilization, but before the sperm can enter the ovum, it must somehow be modified to contain only one centriole. The two hypotheses differ in the way the zygote obtains the second centriole.
Reduction Theory
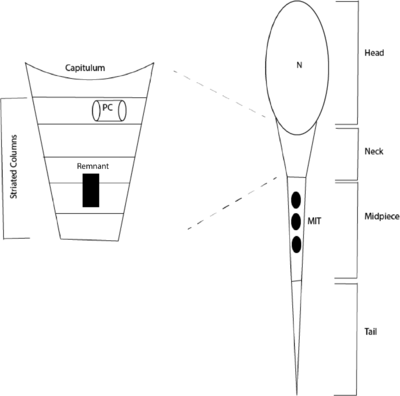
The currently accepted theory is that during spermeogenesis the sperm cells loose their distal centriole. It is hypothesized that the centriole leaves a remnant that is incapable of forming a centrosome, and cannot function after fertilization. This is visualized by Figure 2, depicting a sperm cell that has a single, functional centriole in the striated column. If this hypothesis is true, then in order for the zygote to have the two necessary centrioles, then the proximal centriole must duplicate before each of these centrioles could duplicate again, forming the necessary four centrioles. As no known mechanism for the duplication of the sperm's single centriole exists once it enters the ovum, another hypothesis explains this phenomenon.
A New Hypothesis: Remodeling Theory

Remodeling theory provides an alternative mechanism by which a sperm cell will acquire the two centrioles required to bring the sperm and egg's nuclei together. The basis for the theory is that rather than rendering the distal centriole dysfunctional after maturation, the centriole is remodeled into an atypical centriole. Figure 3 visualizes this hypothesis.
Initial studies in insects preformed by Dr. Avidor-Reiss and his team have led to the discovery of a second, functional centriole that has an atypical structure. This was achieved by staining insect sperm with centriole protein antibodies, and tagging the protein. The results suggested the presence of two centrioles between the sperm head and tail.
This recently discovered centriole is unlike the typical proximal centriole seen below the head of the sperm. Rather, this new distal centriole is a remodeled counter part, and possesses three rod structures in place of a traditional nine-fold design.
The working theory is that this modified centriole is capable of forming a centrosome with the proximal centriole in order to pull the zygotic nuclei together during fertilization.
Centriole Inheritance During Fertilization after Remodeling

During fertilization, the whole sperm enters the ovum. Once inside, the ovum releases more polar bodies, and the male sperm separates the tail from the nucleus. The sperm's nucleus will combine during cleavage with the egg's pronucleus with the assistance of astral microtubules that project from the sperm's proximal/ distal centriole centrosome. This centrosome formation and organization is made possible my the existence of the second, atypical centriole as described by the remodeling theory. Next, the sperm's proximal and distal centrioles (distal still with the tail attached) form pro-nuclei, allowing for the paternal centrioles to form a centrosome with their respective procentriole. These two centrosomes will lead cell division of the embryo by ensuring genetic material and cellular contents are evenly distributed during cleavage.
Further Reading
- Avidor-Reiss, T., Khire, A., Fishman, E. L. & Jo, K. H. Atypical centrioles during sexual reproduction. Front Cell Dev Biol 3, 21, doi:10.3389/fcell.2015.00021 (2015).
- Wilson, Patricia G. “Centriole Inheritance.” National Center for Biotechnology Information, Landes Bioscience, 2008, www.ncbi.nlm.nih.gov/pmc/articles/PMC2634415/.6.
Work Cited:
- ↑ 1.0 1.1 Hirono, Masafumi. “Cartwheel Assembly .” Philosophical Transactions of the Royal Society B: Biological Sciences, The Royal Society, 5 Sept. 2014, www.ncbi.nlm.nih.gov/pmc/articles/PMC4113102/.
- ↑ Sathananthan, A, et al. “The Sperm Centriole: Its Inheritance, Replication and Perpetuation in Early Human Embryos.” OxfordAcademic Journals , Eshre, 1 Feb. 1996,academic.oup.com/humrep/article/11/2/345/606070
- ↑ Azimzadeh, J, et al. “hPOC5 Is a Centrin-Binding Protein Required for Assembly of Full-Length Centrioles.” The Journal of Cell Biology., U.S. National Library of Medicine, 6 Apr. 2009, www.ncbi.nlm.nih.gov/pubmed/19349582/.
- ↑ “Centrin-1.” Centrin-1 - Homo Sapiens (Human), European Bioinformatics Institute Protein Information Resource SIB Swiss Institute of Bioinformatics, 28 Feb. 2018, www.uniprot.org/uniprot/Q12798.
- ↑ Keller, Lani C., et al. “Molecular Architecture of the Centriole Proteome.” Molecular Biology of the Cell, The American Society for Cell Biology, 15 Feb. 2009, www.ncbi.nlm.nih.gov/pmc/articles/PMC2642750/.
- ↑ European Bioinformatics InstituteProtein Information ResourceSIB Swiss Institute of Bioinformatics. “Serine/Threonine-Protein Kinase PLK4.” UniProt, UniProt, 28 Mar. 2018, www.uniprot.org/uniprot/O00444.