Metacommunity ecology (Biodiversity)
Metacommunity ecology
Populations and communities are often patchily distributed across a landscape. Metacommunity ecology refers to the study of the processes involved in “assembly” (gain and loss of species) and maintenance of these patchy communities.Traditional community ecology views communities as the product of local interactions among organisms such as competition for resources. Metacommunity ecology adds to this view by considering communities as the product of these interactions plus larger-scale dynamics created by the movement of individuals among separate communities, often referred to as “patches”. Thus, a high-diversity (Species diversity) habitat, which maintains high population sizes and many co-occurring species, likely results in increased diversity in nearby poorer patches by supplementing populations that would otherwise go extinct. Therefore, metacommuity ecology views community diversity patterns in the context of the larger region.
According to the seminal paper by Leibold et al. there are four main metacommunity processes potentially driving patterns of diversity:
1. Patch dynamics: Classic theoretical approaches to understanding community structure and diversity often assume that species occupy identical habitat patches (for example, see metapopulations), and there are two basic assumptions about patch dynamics. The first assumption is that populations have some rate of local extinction, that is, species will be lost periodically as time passes. The second assumption is that populations have limited dispersal ability (that is, they cannot colonize every patch simultaneously). Given these two assumptions, patterns of diversity are a product of the rates at which species disperse out of patches, which varies among species. Thus, the trade-off between colonization and competitive abilities becomes an important mechanism for understanding coexistence patterns. That is, as long as superior competitors have limited dispersal abilities, their inferior competitors who have better dispersal abilities can disperse to and occupy habitat patches that the superior competitors have not yet reached.
2. Species sorting: Species sorting views landscapes as sets of habitat patches that have differing environments and where the communities on those patches include the species best suited to local conditions. Under this process, the outcomes of interactions (especially competitive exclusion) are thought to occur quickly relative to the rate at which species disperse, thus producing fixed local communities, that is, those with predictable types of species. Further, being part of a large metacommunity with many habitat types means that local species composition can rapidly respond to changes in the environment within local patches because there are many potential species to colonize different types of patches.
3. Mass effects: Mass effects occur when local interactions operate slowly relative to the speed at which species disperse. Thus, local patches maintain populations that would otherwise go extinct without immigration from elsewhere supplementing their numbers. This means two things: first, dispersal is generally thought to increase habitat diversity; second, this effect varies with changes in the rate of dispersal.
4. Neutral dynamics: This final process starts with the assumption that all species are identical in terms of birth and death rates and competitive ability, such that changes in diversity patterns follow random chance. Under these conditions, when local communities are viewed in isolation, local species richness eventually drops to a single species because of the accumulation of random extinctions. But by linking local patches to a larger metacommunity, realistic diversity patterns can be maintained even in the face of such random fluctuations, and a balance between colonization and extinction is struck. Although the assumption of species equality is unrealistic, it allows exploration of what sort of patterns emerge in the absence of differences among species, purely as a result of spatial structure in the environment.
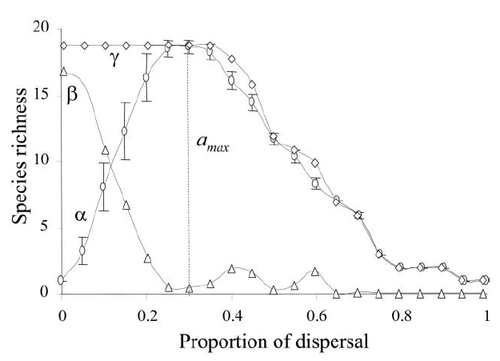
In order to fully understand how different mechanisms of species coexistence operate, metacommunity diversity is usually parsed into three components: local patch diversity, among-patch diversity (which measures how different patch communities are from one another), and total metacommunity diversity. While there are several ways to calculate these diversities, Lande’s additive partitioning scheme, where local diversity and among-patch diversity add up to total metacommunity diversity, is usually used for metacommunity studies. The relative magnitude of the local and among-patch portions of metacommunity diversity inform our understanding of the potential mechanisms affecting species coexistence (Fig. 1). High local and low among-patch diversity may indicate that within-habitat interactions (such as niche partitioning) are the main drivers of diversity, whereas with high among-patch diversity, one may infer that environmental differences and limited dispersal create a mosaic of the places where species occur.
The term metacommunity is usually used to refer to a set of processes that generate diversity patterns, rather than for a defined region or discrete set of patches. However, this dichotomy between conceptual and spatial definitions has lead to two types of metacommunity studies. First, studies that examine diversity patterns at the scales (local, among-patch and metacommunity) where every patch is followed are routinely done with theoretical/computational and aquatic laboratory studies. Here specific variables are manipulated (such as dispersal or extinction) that allow researchers to test the effects of specific processes on diversity patterns. In the second type of study, researchers examine spatial variation among real-world communities and use statistical methods to parse out the different processes by accounting for the effects of habitat and environmental differences or spatial distances among patches.
See also
References and further reading
- Burns K.C, & Neufeld C.J. (2009) Plant extinction dynamics in an insular metacommunity. Oikos, 118: 191-198
- Cadotte M.W. & Fukami T. (2005) Dispersal, spatial scale, and species diversity in a hierarchically structured experimental landscape. Ecology Letters, 8, 548-557
- Cottenie K. (2005) Integrating environmental and spatial processes in ecological community dynamics. Ecology Letters, 8, 1175-1182
- Holyoak M., Leibold M.A. & Holt R.D. (2005) Metacommunities: Spatial Dynamics and Ecological Communities. University of Chicago Press, Chicago, IL.
- Horn H.S. & Macarthur R.H. (1972) Competition among Fugitive Species in a Harlequin Environment. Ecology, 53, 749-752
- Lande R. (1996) Statistics and partitioning of species diversity, and similarity among multiple communities. Oikos, 76, 5-13
- Leibold M.A., Holyoak M., Mouquet N., Amarasekare P., Chase J.M., Hoopes M.F., Holt R.D., Shurin J.D., Law R., Tilman D., Loreau M. & Gonzalez A. (2004) The metacommunity concept: a framework for multi-scale community ecology. Ecology Letters, 7, 601-613
- Mouquet N. & Loreau M. (2003) Community patterns in source-sink metacommunities. American Naturalist, 162, 544-557
- Tilman D. & Kareiva P. (1997) The role of space in population dynamics and interspecific interactions. Princeton University Press, Princeton, NJ.
- Whittaker R.H. (1965) Dominance and diversity in land plant communities. Science, 147, 250-260