Ecophylogenetics
Ecophylogenetics is a field of study that uses information about the evolutionary relationships among species (phylogenetics) to test hypotheses about how ecological communities are assembled and about the potential outcomes of species interactions. A central problem in community ecology is what determines which and how many species live together in a community. The coexistence of species within communities is often predicated on either of two opposing mechanisms: environmental filtering, whereby species co-occur in communities based on shared tolerances and requirements, and niche differentiation, whereby species coexist because they occupy different niches, which reduces competition. The first mechanism (environmental filtering) predicts that similar species should coexist in the same habitats, whereas the second (niche differentiation) posits the opposite pattern—that coexisting species should be different from one another.
Distinguishing between these two mechanisms requires characterizing the niches of species in a community. Defining and measuring these niche differences among species has proven to be rather difficult because of the large number of characteristics that could be measured and the multifaceted ways in which species could be different from one another. For example, plant ecologists could measure leaf size, thickness, lobiness, chemical content, hairiness, density and the rate of water loss and species differences could be evaluated from any one of these measures or any number simultaneously. Another important consideration is that species similarities and differences are products of their evolutionary history, such that the more history two species share (that is, the more recently they diverged from a common ancestor), the more similar they are likely to be as compared to species with whom they shared more distant common ancestor. Thus, distinguishing the mechanisms involved in community assembly requires knowing the evolutionary history of a group of species, which is represented by a special type of graph called a phylogeny, essentially a family tree of the species (Fig. 1).
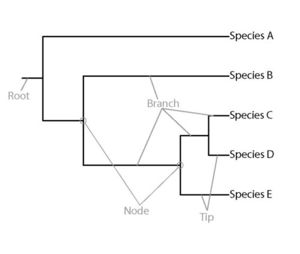
Longer branch lengths (indicative of the amount of evolution since the two diverged) connecting a particular species to other species in the tree generally means greater uniqueness of traits or occupation of a more unique niche. Similarly, fewer nodes (branching points) along the branches connecting two species to the root of the phylogeny mean those species shares less evolutionary history. By measuring the lengths of phylogenetic branches connecting species together, researchers can estimate the potential uniqueness of species.
A number of scientists, dating back to Charles Darwin, thought both about species origins and the evolution of traits and niches while also thinking about why species occur where they do. Campbell Webb, working in tropical forests, was the first to explicitly use phylogenetic information to examine and attempt to explain patterns of species co-occurrence. He tested whether species within communities were more or less closely related to other community members then predicted from randomly generated communities. This approach uses patterns of phylogenetic relatedness to potentially exclude environmental filtering or niche differentiation as plausible mechanisms for how particular community structures developed. (Fig. 2).
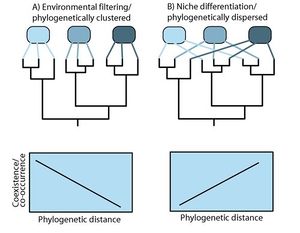
We should expect that environmental filtering determines the structure of ecological communities because an organism’s basic niche requirements and tolerances evolve relatively slowly and thus closely related species are generally very similar in both appearance and ecology. Jeannine Cavender-Bares pioneered research into untangling the opposing forces of environmental filtering and niche differentiation for detecting and understanding the phylogenetic influences on community patterns. With colleagues, she showed that the relative strength of filtering versus differentiation depends on the size of the land area and the number of species being observed. These researchers showed that at larger spatial scales, such as landscapes, environmental filtering appears a better explanation for patterns of species coexistence, whereas niche differentiation can be more important at smaller scales. A similar pattern emerged when they narrowed their taxonomic focus: they found that when they considered all the plant species in communities, the communities appeared to contain more closely-related species, whereas when they examined more specific groups, such as the oaks, they found that communities actually consisted of less closely related species. There are now several such examples of this scale-dependency in the mechanisms operating during community assembly. Helmus, Matthew|Mathew Helmus]] and colleagues showed that environmental filtering and differentiation operate simultaneously in fish communities, as have Lovette and Hochachka for birds across North America. Thus, ecophylogenetic analysis of communities gives investigators useful information about the plausible mechanisms producing patterns of community coexistence.
While the phylogenetic relationships among species ultimately represent trait and niche differences, the information required to adequately define total trait or niche differences among species is often tedious and problematic to collect. But knowing how the similarities and differences among species affect ecological processes is also critically important for conservation decisions. For example, the knowledge of which species are likely to be affected by environmental changes and which can be introduced into new habitats is central for making sensible management decisions. Using the evolutionary history among groups of species may be the easiest way to estimate and predict the similarities and differences in responses of species to management and conservation decisions. For example, Strauss and colleagues showed that highly invasive annual grasses in California tend to be less related to the native community than noninvasive species that are introduced. Further, Cadotte and colleagues found that the local extinction of more distinct plant species caused a greater decline in community productivity than extinction of less distinct species. Both of these example studies illustrate a starting point for how to evaluate species differences and their potential importance in ecology and conservation biology.
Further reading
- Cadotte, M. W., B. J. Cardinale, and T. H. Oakley. 2008. Evolutionary history and the effect of biodiversity on plant productivity. Proceedings of the National Academy of Science, USA: In press.
- Cavender-Bares, J., A. Keen, and B. Miles. 2006. Phylogenetic structure of floridian plant communities depends on taxonomic and spatial scale. Ecology 87:S109-S122.
- Cavender-Bares, J., and A. Wilczek. 2003. Integrating micro- and macroevolutionary processes in community ecology. Ecology 84:592-597.
- Felsenstein, J. 1985. Phylogenies and the comparative method. American Naturalist 125:1-15.
- Harvey, P. H., and M. Pagel. 1991, The comparative method in evolutionary biology. Oxford, Oxford University Press.
- Helmus, M. R., K. Savage, M. W. Diebel, J. T. Maxted, and A. R. Ives. 2007. Separating the determinants of phylogenetic community structure. Ecology Letters 10:917-925.
- Lovette, I. J., and W. M. Hochachka. 2006. Simultaneous effects of phylogenetic niche conservatism and competition on avian community structure. Ecology 87:S14-S28.
- Stephens, P. R., and J. J. Wiens. 2004. Convergence, Divergence, and Homogenization in the Ecological Structure of Emydid Turtle Communities: The Effects of Phylogeny and Dispersal. American Naturalist 164:244-254.
- Strauss, S. Y., C. O. Webb, and N. Salamin. 2006. Exotic taxa less related to native species are more invasive. Proceedings of the National Academy of Sciences of the United States of America 103:5841-5845.
- Webb, C. O. 2000. Exploring the phylogenetic structure of ecological communities: an example for rain forest trees. American Naturalist 156:145-155.
- Webb, C. O., D. D. Ackerly, M. A. McPeek, and M. J. Donoghue. 2002. Phylogenies and community ecology. Annual Review of Ecology and Systematics 33:475-505.