Mutation

Mutations in carrots have produced overt color distinctions. USDA
 Irradiation of pineapple plants is being conducted to induce mutations
Irradiation of pineapple plants is being conducted to induce mutations
in search of higher productivity and thorn reduction. PNRI A mutation is an alteration of the genomic sequence of DNA molecules. Mutations may cause altered function in the affected organism, or may cause alteration of inherited traits of offspring, or no change at all. In the case of viruses the mutation may apply to DNA or RNA molecules, depending upon the viral species.Mutation an important driver of evolution. Mutations are generated by a variety of causes: some external to the organism and some internal. External causes include impingement of some forms of electromagnetic radiation, such as x-rays or ultraviolet radiation, and may also include exposure to mutagenic chemicals or viral infection. Mutation processes not induced by any external forces include genomic replication errors, transcription errors, code sequence transpositions and an immune system response known as somatic hypermutation.
The physiological consequences of mutation are quite variable, ranging from death of a single cell, carcinoma to heritable genetic outcome - or they a mutation may be repaired through DNA repair mechanisms. Mutations in reproductive (germline) cells of multicellular organisms generally produce inheritable consequences, while mutation within other tissues (somatic) ordinarily only have outcomes affecting the individual in which the mutation occurs.
Contents
- 1 Consequences of mutation
- 2 Types of mutations
- 3 Role in natural selection
- 4 Role in research and medicine
- 5 The molecular view
- 5.1 Single nucleotide polymorphism
- 5.2 Nucleotide deletion
- 5.3 Frameshift mutation
- 5.4 Simple variable number of tandem repeat polymorphism
- 5.5 Inversion polymorphism
- 5.6 Large scale sequence variants
- 5.7 Fourfold degenerate sites
- 5.8 Twofold degenerate sites
- 5.9 Transposon mutation
- 5.10 Deamination
- 5.11 Alkylation
- 6 Chemical mutagens
- 7 The numbers game
- 8 References
Consequences of mutation
 Drosophila melanogaster offspring with variation in eye and
Drosophila melanogaster offspring with variation in eye and
body color; this is a model organism for studying mutations.
Depending upon the molecular nature of the mutation, and depending on the locus in specialized cells versus reproductive cells, the consequences of mutation range from insignificant to catastrophic; moreover, in the majority of mutation instances which represent non-coding occurrences, there is usually no immediate impact upon cellular function or heritable trait change; however, there may be latent mutations that reside in the non-coding segments that can be important in gene expression for future generations.
Where mutations occur in coding sequences within non-reproductive cells, the impact to an organism may be a minor alteration in metabolic efficiency (either beneficial or deleterious); in extreme cases cell death may occur, when the metabolic change is adverse and highly consequential. Other adverse outcomes may produce a cell that has abnormal morphology, combined with exceptional replication robustness; these circumstances are usually considered carcinoma events, where the mutated cells take over the native tissue and may invade other body organs. Ultraviolet or X-ray radiation induced mutations may either cleave the DNA molecule entirely (in which case reproduction is not likely to occur), or else cause pointwise mutations (which outcome will sometimes lead to formation of a cancerous cell).
In the instances that genetic mutations result in morphologically distinguishable offspring, there are potentially significant implications for the species evolution and survival. Some research has suggested that exposure of amphibians and other fauna to mutagenic pesticides can lead to dramatic changes in organism morphology and corresponding fitness reduction. These outcomes are often called deformities, and their consequences will be elaborated in the discussion of natural selection below.
Types of mutations
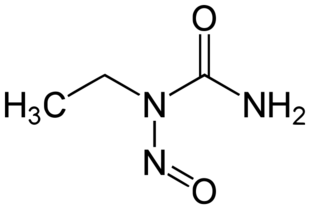 Ethylnitrosourea, an alkylating mutagenic agent. There are a number of distinct types of mutation:
Ethylnitrosourea, an alkylating mutagenic agent. There are a number of distinct types of mutation:
- Mutation in non-coding genomic sequences, which have no known effect upon organism traits or metabolism
- Beneficial mutations that allow inherited traits of greater fitness or reproductive success
- Adverse (including some carcinogenic) mutations that allow inherited traits of reduced fitness or reproductive success)
- Non-heritable mitochondrial mutations that provide different coding instructions for protein assembly
- Lethal or carcinogenic mutations that threaten the life of the organism, but are not heritable
- Mutations that diversify the genomic character and may assist in future generation adaptability
Role in natural selection
Mutations are fundamental to the phenomena of genetic variability and natural selection. Such processes must be viewed as inheritance transitions spanning at least hundreds or thousands of reproductive generations. Mutations may produce viable offspring that have differences in traits from parent stock. After this molecular clock has ticked for thousands of generations, there may be a panoply of genomic mutations that represent a richness of characteristics that exhibit (usually) subtle differences in the organism of interest. Mutations which are favorable to species fitness, survival and reproduction are preferentially propagated to the successive generation, since higher reproductive success is expected for individuals holding these beneficial mutations. Conversely, individuals who have adverse mutations are discriminated upon in their propagation and fecundity.
Role in research and medicine
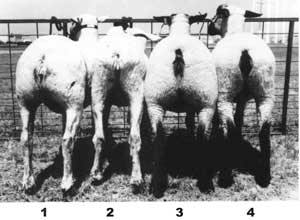 Sheep 1 and 3 are offspring of mutants having the callipyge (robust rump) syndrome. USDA Mutations have been noted and used in agricultural research for centuries. Dedicated crop and animal scientists have utilized random mutations for breeding of more productive species as well as more disease resistant species to feed the rapidly expanding human population.
Sheep 1 and 3 are offspring of mutants having the callipyge (robust rump) syndrome. USDA Mutations have been noted and used in agricultural research for centuries. Dedicated crop and animal scientists have utilized random mutations for breeding of more productive species as well as more disease resistant species to feed the rapidly expanding human population.
In some cases these efforts have led to overzealous production of narrow breeding lines that endanger the gene pool of a diverse seed or breeding stock, and amplify the adverse effects of monoculture farming systems.
In more modern times scientists are applying knowledge of mutations to combat cancer, specifically targeting tumors with agents that can cause such damage to cancer cells that they are preferentially destroyed. These techniques include radiation as well as chemical mutagens; while many such efforts offer great promise, the spatial targeting of these measures are critical hurdles in defining effective portocols.
The molecular view
Most altered DNA or mutations arise from substitution or deletion of one or more nucleotides. If the nucleotide affected represents a coding portion of the DNA molecule, there results an alteration of amino acid selection in the instructions within the cell that construct essential proteins required for cell functioning. If the mutation occurs in a non-coding portion of the DNA molecule, there is no impact to the organism, although, if the non-coding mutation occurs in the egg or sperm cells, the altered DNA sequence may be inherited, with an expectation that there will be no impact to traits of successive generations.  Double helix DNA model
Double helix DNA model
Single nucleotide polymorphism
The simplest form of replication error mutation is the substitution of a single nucleotide; most commonly there are only two different alleles that can result from this type of polymorphism due to constraints of stereochemistry and base pair bonding. Only about 1.5 percent of these single nucleotide polymorphisms occur in the coding portions of the DNA molecule. In general the non-coding substitution mutations do not result in genotype expression, except in some cases where the non-coding sections influence instructions for cellular metabolic regulation.
Nucleotide deletion
When a single base is deleted in a replication process, it is termed a deletion mutation. This form of mutation is very difficult for the cellular DNA apparati to repair, and can lead to a frameshift mutation described below.
Frameshift mutation
In non-coding sequences, it is relatively common to observe insertion/deletion mutations where one or more nucleotides is involved; however, such mutations are rare in coding sequences, probably due to the fact that resulting codons often interrupt the replication process itself. Thus, such mutations may not propagate to successive generations. Such nucleotide substitutions which are numerically not divisable by three, and fail to replicate are termed frameshift mutations.
Simple variable number of tandem repeat polymorphism
This phenomenon is characterized by Alleles containing tandemly repeated runs of a simple coding sequence. In the case of microsatellites, or simple sequence repeats, the sequence may be a single or several base pairs long, but the array length may extend from about ten to more than 100 nucleotides. An alternate form of minisatellites, the total array may span hundred of nucleotides, and the loci frequently possess multiple alleles.
Inversion polymorphism
There are a number of examples of high sequence homology within portions of the genome. For example, within the euchromatic, or genetically active portion of the human genome, there is a very long repetitive sequence, where inversion polymorphisms commonly occur; moreover, this specific portion of the genome is one common to the coding for all primate animals. This very long high homology sequence is predisposed to large scale deletion, duplication and inversion polymorphism. In the case of inversion polymorphisms there is a low incidence of deleterious or disease outcome, which adverse outcomes are often associated with the duplication and deletion scenarios.
Large scale sequence variants
Sometimes very large scale mutations occur altering hundreds to thousands of consecutive base pairs in non-coding portions of the genome. These occurrences are alternatively termed chromosomal polymorphisms, and they can often be detected by conventional cytogenetic methods because of their extraordinary size; conversely, due to the occurrence in non-coding portions of the genome, they have little or no impact upon cellular metabolism or trait alteration.
Fourfold degenerate sites
Some gene loci are in codon positions where a nucleotide substitution does not affect the specifications for codon translation. In these cases there is no effective outcome of a nucleotide substitution mutation. The loci of such substitutions are termed fourfold degenerate sites, and this type of mutation comprises approximately sixteen percent of nucleotide substitution mutations.
Twofold degenerate sites
This form of coding sequence replacement that affects either the first or third base pair in the codon. In these substitutions only two of three of the substituted will code for a new amino acid. Replacement by a base that does not affect amino acid coding is sometimes called a silent mutation, since there is no functional change ensuing from a replacement base, where the resulting sequence codes for the original amino acid of the codon.
Transposon mutation
This process permits a gene to be transferred to the chromosome of a host organism, altering or interrupting functionality of a gene and inducing a mutation.
Deamination
Hydrolysis may transform a normal base to an atypical one that has a keto group instead of the original amine group; for example, cytosine can be altered in this fashion to produce uracil. This type of mutation is considered a pointwise spontaneous event since no external forces are involved; it is a mutation type that is most readily corrected by DNA cellular repair mechanisms.
Alkylation
Mutation of nucleotide bases may occur by presence of certain mutagenic chemicals. The process of alkylation is an example of such an incursion by a foreign chemical into the chromosomal area. A specific mutagen known to high a high frequency of mutations is nitrosourea, which ethylates one of the bases, usually thymine.
Chemical mutagens
 3-chloro-4-(dichloromethyl)-5-hydroxy-5H-furan-2-one, a mutagen present in many water systems.
3-chloro-4-(dichloromethyl)-5-hydroxy-5H-furan-2-one, a mutagen present in many water systems.
There are a number of chemicals implicated in mutation phenomena. Some are indigenous to the organisms naturally, and some are taken into the cells via environmental exposure. Chief among these mutagens are:
- Reactive oxygen species, a set of oxidative agents that are almost always present in living cells, but whose elevated concentrations pose risk to DNA and its replication
- Alkylation molecules, capable of substituting a methyl, ethyl or other alkyl modification to a base
- Certain aromatic compounds, including derivatives of benzene, pyrene and anthrocene
- Nitrous acid, which can convert an amine group of cytocine or adenine to a diazo group; nitrous oxide can also be classed as an indigenous DNA crosslinking element.
- Intercalating agents, which can insert themselves within the DNA double helix and interfere with normal replication (examples include berberine, thalidomide and proflavine)
- Molecules that can substitute for a nucleotide, such as 5-bromouracil
- Molecules that can covalently bond to a segment of DNA (These chemicals are often carcinogenic; acetaldehyde is an example molecule.)
The numbers game
From a statistical standpoint, it is easy to understand why replication mutations can occur. During a human lifespan, for example, approximately 1017 cell divisions occur; compounded with this staggering number of instances where replication errors can take place, one must remember that each chromosomal reproduction needs about six billion matchups of base pairs. These numerical aspects are not only sobering, but also underscore the necessity of DNA repair mechanisms at work in the replication process.
In fact, the natural replication process that employs DNA polymerase is documented to have about one error (or mutation event) in every ten trillion base pair selections; while that uncorrected number is impressive, it would still lead to an unacceptable rate of mutations, were it not for the DNA repair capabilities inherent in all organisms.
References
- George C.Williams. 1966. Adaptation and natural selection: a critique of some current evolutionary thought. Princeton Science Library. "Evolutionary adaptation is a phenomenon of pervasive importance in biology".
- T. Strachan and Andrew P. Read. 2004. Human Molecular Genetics. Garland Science. 674 pages
- S.Hitotsumachi, D.A.Carpenter and W.L.Russell. 1985. Dose-Repetition Increases the Mutagenic Effectiveness of N-ethyl-N-nitrosourea in Mouse Spermatogonia. Proc. Natl. Acad. Sci. USA 82, 6619-6621
- J.Bertram. 2000. The molecular biology of cancer. Mol. Aspects Med. 21 (6): 167–223.
- Daniel L.Hartl and Elizabeth W.Jones. 2009. Genetics: analysis of genes and genomes. Jones & Bartlett. 763 pages
- A.D.Richards and A.Rodgers. 2007. Synthetic metallomolecules as agents for the control of DNA structure. Chem. Soc. Rev. 36: 471–483.