Mollusca

Clams, bivalve molluscs.
Contents
- 1 Introduction Molluscs are soft-bodied invertebrates whose main features are a foot for locomotion, a mantle which secretes a hard calcium shell, a general head region, and a stomach area for digestion. Molluscs are very diverse with about 110,000 species distributed in terrestrial, freshwater and marine environments. The Gastropoda (snails) include the largest number of species comprising over three quarters of species in this phylum. The Bivalva (clams/oysters/mussels) is less diverse but also a very important group. (Mollusca)
Introduction Molluscs are soft-bodied invertebrates whose main features are a foot for locomotion, a mantle which secretes a hard calcium shell, a general head region, and a stomach area for digestion. Molluscs are very diverse with about 110,000 species distributed in terrestrial, freshwater and marine environments. The Gastropoda (snails) include the largest number of species comprising over three quarters of species in this phylum. The Bivalva (clams/oysters/mussels) is less diverse but also a very important group. (Mollusca)
Bivalva
Introduction (Source: Biodiversity Institute of Ontario (Mollusca) )
Bivalves are the clams and mussels of our freshwater lakes. The word Bivalva (Bi= "two", valva="shells") is a misnomer. Actually, there is only a single shell secreted with a left and right lobe. Many people tend to think that one half of a clam shell is the top and the other is the bottom. Actually, these animals are laterally flattened– there is a shell on the lefthand side and the right hand side. The mouth (incurrent canal) and anus (outcurrent canal) are located at the anterior end, and while the foot extends out the posterior.
Morphology
Adult Morphology
The mantle of all bivalves consists of three folds of tissue. The outer fold secretes the shell, the middle fold contains sensors, and the inner fold contains the radial and circular muscles.
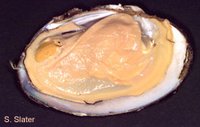 (Source: Biodiversity Institute of Ontario)
(Source: Biodiversity Institute of Ontario) The mantle is attached to the cell at certain muscle insertion points. Otherwise there is a narrow gap between the two structures. Sometimes a parasite or grain of sand lodges in this gap and is coated with secretions. The foreign object may be either plastered to the shell to form a protrusion or it may be absorbed into the mantle where a spherical pearl forms from multiple layers of secretion.
The foot is flattened and can be extended between the two valves using blood pressure, and maneuvered by muscles called pedal protractors and retractors. As it is spade-shaped, the foot is an ideal tool for digging into the bottom sediments.
Since the entire body of a bivalve is ordinarily enclosed inside its shell, it does not come into contact with the outside environment. As a result bivalves lack a distinct head region with the sensory structures that the heads of other animals have such as hearing, taste, and sight. On the middle fold of mantle, along the outer edge of each shell, there are photoreceptor cells that detect differences in light intensity, although they cannot form images. If the shadow of a predator falls on a bivalve, it reacts by closing its shell.
Larval Morphology
 (Source: Biodiversity Institute of Ontario)
(Source: Biodiversity Institute of Ontario) Although bivalve species with free-swimming larvae are abundant in marine environments, they are much less common in freshwater. Zebra mussels have a free-swimming veliger larva. Its structure is very different from the ecto-parasitic glochida larvae that are typical of all freshwater clams. Glochidia are separated into three different types based on their host attachment structures.
Hooked glochidia: are found only in species of the subfamily Anodontina. On each of the shell margins furthest from the hinge they bear a large incurved hook that is inlaid with sharp spines. These larvae are adapted to cut through fin and scales in order to attach to their fish host.
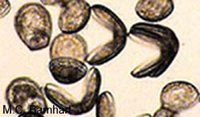 (Source: Biodiversity Institute of Ontario)
(Source: Biodiversity Institute of Ontario) Non-hooked glochidia: do not have large hooks, but their inner shell margins have fine sharp spines. When the shell "bites down" these smaller spines attach well to gill filaments.
Axehead glochidia: are found only in the genus Proptera and have a left and right shell lobe that flares outward giving the appearance of an axe head. On most species there are small hook-like structures on each corner. No one is sure where these parasitic larvae attach to their host.
Adult shell morphology
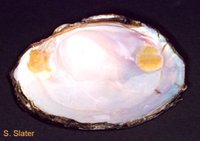 (Source: Biodiversity Institute of Ontario)
(Source: Biodiversity Institute of Ontario) Like snails, the shells of bivalves have a dark, leathery outer layer of protein that helps protect the inner calcium carbonate later from abrasion and calcium loss. However, the shells of bivalves have two inner layers. The only difference between these two layers is the arrangement of the calcium carbonate crystals is skewed 90 degrees in the middle layer. The middle calcified layer is a dull white, while the inner layer is a shiny pearl colour. This innermost portion is constantly secreted by the mantle and is what gives shells their rigidity.
Metabolism
Bivalves have an open circulatory system. A system of this type does not have thin walled capillaries through which nutrients and oxygen can diffuse to the cells that need them. Instead the blood (or haemolymph) flows into open cavities and bathes a large surface area of cells, passing on the needed materials directly, before returning to the heart. Bivalves do have veins and arteries that branch out, but they simply empty into open cavities instead of the capillary beds.
When bivalves are ready to excrete their undigested wastes, they package it up in mucous before ejection. This is an important step since the inhalant and exhalant canals are located right beside one another. Since the faeces are packaged and heavy when released, they avoid being re-ingested.
Reproduction/Development
The majority of freshwater bivalves are hermaphroditic and produce mature sperm and eggs at the same time, often leading to self fertilization. Egg and sperm formation is triggered when water temperature rises above a critical mark. The few species that have separate sexes display sexual dimorphism: the gills of the female enlarge to contain the developing larvae. The males have no copulatory organs since the adults are sessile and unable to make contact with one another.
 (Source: Biodiversity Institute of Ontario)
(Source: Biodiversity Institute of Ontario) The invading zebra mussel, Dreissena polymorpha, has a different reproductive system than North American bivalves. Zebra mussels shed both sperm and eggs into the water, and its spherical eggs are much smaller than other freshwater species at a diameter of 40-70 micrometres. Since these eggs are so small and carry little nutrition for the developing embryo, the larvae hatch out quickly. These veliger larvae are free-swimming unlike those of other freshwater bivalves. These larvae filter-feed in the water column before settling to the benthos to transform into adults. North American freshwater clams release sperm into the water column where it is ingested by the inhalent siphon of a neighboring female clam. The eggs, having already been transferred from the female's gonopore to her gills, await fertiliation as the sperm are passed over the gills. These gill tissues are modified into a gill marsupium, which protects and aerates the eggs until fertilization. Freshwater bivalve eggs are larger and have more yolk than their marine counterparts. The yolk serves to feed the developing embryo and larva until it attaches to a host. The larvae pass through the veliger stage, while they are still in the marsupium and are only released after they transform into the glochidial stage. At this time, the larvae are shot into the water from the exhalent siphon. The glochidia then attach to a vertebrate host in which the encyst and transform, protected and fed by the vertebrate tissue, into an adult.
Ecology
Adult ecology
Adult bivalves utilize gills to extract oxygen from the water, as do many other aquatic organisms. However, these gills are also the main structure used to acquire food. All Canadian bivalves are filter feeders; they take in water and any food particles that are suspended in it, with the inhalant siphon. This water passes over the gills where two things happen before the water is returned to the environment by the exhalant siphon. Oxygen is removed from the water and carbon dioxide is released into it. At the same time the particles suspended in the water (microorganisms, detritus, algae) are stuck to the gill filaments by the mucous that is secreted by the gills. Each of the two gills possesses a food groove at its base, which collects the tiny particles that have been captured. Particles that stick to the gills are sorted into food and non-food items. Those classified as food are transported to the food groove by cilia. At the end of this chain are the labial palps, a pair of which lie on either side of the mouth, that collect the food and push it into the mouth.
There are more bivalve species in North American waters than in any other freshwater bodies in the world: 260 indigenous species and 6 introduced species. Bivalves belonging to the Superfamily Unionacea are the most prevalent, aided in distribution by their larvae which are parasites on fish.
Most Canadian bivalves burrow using their fleshy foot, that extends from the posterior end between the shell halves, to dig into the substrate using circular muscles that cause it to extend. The foot is then anchored in the sand/organic matter as haemolymph is pumped into the end of the foot. (To get a better understanding of how this is done, see the circulation section for this animal). A burst of water is then forced from the crack between the shell. This force loosens the substrate and the clam pulls itself into the gap. They pull themselves deeper and deeper in this fashion until they are almost completely buried with only their anterior end sticking out.
Larval ecology
When glochidia larvae attach to their host fish they damage it by digging into its flesh. The immune system of the fish attempts to heal the body by sending blood and mucous to the site of the invasion. Unfortunately for the host, this is exactly what the glochidia feed upon. Adult fish are not grievously harmed by the attachment, but smaller fish are often overcome by an infection at the attachment site. After feeding for 2 to 36 hours, the glochidia encyst. During this period they do not feed, are protected by the tissues of the host, and begin their transformation into the adult form. This phase typically lasts for 10 to 30 days. When transformation is complete, the clam emerges from the cyst and drops to the lake bed, where it finishes its growth.
Most glochidia do not need structures for locomotion. Like all kids they rely on a parent to give them a lift wherever they want to go. And like all good parents, adults clams want to be sure that their kids have everything they need to grow into adults so they can finally move out of the house. In the case of the bivalves this means attracting a suitable fish host for their offspring. Different species use different tactics. Some species bind their larvae into mucous threads and release them into the water. These chains of glochidia look amazingly like worms that many fish find tasty. Fish consume these threads and the larvae attach themselves to its gills as the water passes through. Other groups lure the fish closer to home, as adult adults extend their mantle flaps outside their valves so they wave in the currents to give the appearance of bottom dwelling worms. When a fish dives in for a meal, the "worm" retracts and a cloud of glochidia is released.
 (Source: Biodiversity Institute of Ontario)
(Source: Biodiversity Institute of Ontario)  (Source: Biodiversity Institute of Ontario)
(Source: Biodiversity Institute of Ontario) Glochidia larvae have a chemical sense that is triggered by the "scent" of fish blood, mucous, and gill tissue. This sense warns them when there is a fish swimming near enough for attachment. The larvae will not respond to water currents or touch by other organisms. This sense is attuned only to chemical secretions of fish.
Idiosyncratic Inverts
Idiosyncratic inverts I
If there is a drought, bivalves can survive exposure in an air environment. As they lack breathing adaptations for an out of water experience, they simply clam up. This seems to work, as some clams can spend months, even years out of water and still be revived. They have special mechanisms to deal with the obvious problems:
Problem #1: Clams can't process oxygen that isn't dissolved in water.
Solution: Don't use oxygen. Bivalves survive on anaerobic respiration when out of water. All this means is that it sustains all essential metabolic functions without oxygen. This anaerobic metabolism is much slower, and produces less waste.
Problem #2: Metabolic wastes from anaerobic metabolism can build up and poison the clam.
Solution: To avoid this danger, the clam opens slightly to expel the wastes. Vital moisture also escapes, so it can't be done too often or the bivalve will dry out.
Problem #3: Acidity resulting from anaerobic metabolism builds up in the blood (haemolymph).
Solution: In order to buffer these acids, the bivalve diverts calcium ions from its shell to the blood, allowing for a greater storage of the acidic wastes.
When dried-out bivalves are re-submerged in water, they quickly open and circulate as much water through their bodies as they can; flushing out wastes and taking in massive amounts of oxygen. Within 12 hours, things are back to normal.
Idiosyncratic inverts II
Adult bivalves have a unique stomach. After food particles enter their mouth they travel down the esophagus (throat) where they are attached together, by mucous, as a giant chain. The floor of the stomach possesses a sword-like projection of its internal lining. There is a crystalline nail at its tip that reaches all the way to the ceiling of the stomach. This protein crystal is continuously generated by the stomach, and the only reason why it doesn't poke a hole in the top of the stomach is because it is protected by a chitinous (a very hard protein) shield. As the nail grows, it rotates slowly and grinds against the shield. The mucus/food chain that enters the stomach is wound around the nail by this spinning motion. When the food reaches the junction of the nail and the plate, it is ground down by the friction between these two structures. This is how a bivalve "chews" its food. At the same time, chemical digestion is initiated by the enzymes that are secreted through tiny pores in the plate by the cells behind.
Gastropoda
Introduction (Source: Biodiversity Institute of Ontario (Mollusca) 2)
The word Gastropoda is a combination of two Latin words that translate to mean "stomach" and "foot". Canadian freshwater gastropods are divided into two groups: the prosobranchs and the pulmonates.
Pulmonates were originally aquatic until, at some point in history, they invaded land. In doing so they developed a range of adaptations needed to survive in terrestrial habitats. For example, in the water they used ammonia as a vehicle to excrete all their wastes, but changed to a less toxic compound, urea, to facilitate their survival on land. These snails also lost their gills upon transfer to land. When the pulmonates returned to the water they never regained these lost attributes. The prosobranchs, however, never ventured onto land, so their body functions have been continuously adapting to an aquatic environment.
Morphology
Body Morphology
The gastropod body consists of the mantle and the foot. The mantle houses the viscera and gonads which are protected by the calcareous shell. Inside the spiral type shell the mantle extends all the way to the apex, becoming thinner as is ascends. In the pulmonates the mantle cavity has been transformed into a lung, trapping air for underwater respiration. The foot extends out of the shell aperture for locomotion. The muscles in the foot contract in a wavelike motion to pull the snail across the substrate. The indistinct head with sensory structures flows together with the foot and is withdrawn inside the shell if danger threatens.
Freshwater snails have eyes much like those of terrestrial snails. However, instead of being on the end of stalks, they are directly attached to the head region.
Shell Morphology
Snail shells are important for use in systematics, that is deciding who is more closely related to whom. There are three basic shell shapes, all of which have different structural strengths.
Conical: These resemble one half of a clam shell and are restricted to one group of freshwater snails: the limpets.
Planospiral: These snails secrete their shell in ever larger circles. As these snails age their shells get wider and wider.
Spire: These shells look almost like a spiral staircase. The "steps" are widest at the bottom and slowly get smaller until they reach a single point at the top step. There is a central support, called a columella, that runs right through the middle of the shell which gives it strength and provides a point of attachment for the soft parts of the animal.
 (Source: Biodiversity Institute of Ontario)
(Source: Biodiversity Institute of Ontario)  (Source: Biodiversity Institute of Ontario)
(Source: Biodiversity Institute of Ontario)  (Source: Biodiversity Institute of Ontario)
(Source: Biodiversity Institute of Ontario) All three shell types are made of the same material. The inside of the shell is crystalized calcium carbonate mixed with protein. The calcium is absorbed from the snail's food and from the surrounding water. Those snails that live in environments with high amounts of calcium will grow thicker shells than those that do not. When there are only small amounts of calcium available in the water, the snail has to spend much more energy to build its shell, so it only makes the minimum thickness required to survive.
The outside layer of the shell is mostly protein and serves to protect the inner layer from scratches. It also defends the shell from acidic lake water that would cause the calcium carbonate to dissolve thereby weakening or destroying the shell.
Metabolism
This is yet another instance when the prosobranchs and the pulmonates differ. What they use to breathe is also reflected in how these two groups are named. The word branchs, (pronounced "branks") in prosobranchs, means 'gill'. The prefix pulmo in pulmonates refers to a lung. In humans we have a pulmonary system which is the interface between the lungs and the blood which expunges carbon dioxide from and absorbs oxygen into the blood stream.
The prosobranchs possess a very efficient gill system that works the same way as a fish gill. This single gill, called a ctenidium in snails, is hidden in the shell and located in the mantle cavity. Tiny cilia on the gill create a current to direct oxygen-rich water over the gill. The oxygen-poor blood inside the snail is funneled through the gill in the opposite direction of the current created by the cilia. Due to the large gradient in oxygen levels between the water and the blood the oxygen enters into the blood stream which supplies the rest of the snail's body. This is called a counter-current exchange system.
The pulmonates lack a gill; instead the use a pocket formed in the mantle as a primitive lung. This pocket is lined with many tiny blood vessels to make the exchange of oxygen very efficient. Many pulmonates rise to the surface of the water to exchange air. They have a very low metabolism so a breath lasts them a long time. Many species also acquire oxygen through their skin until they rise to the surface. Some groups have developed hemoglobin (the same molecule that is in human blood) to help transport oxygen more efficiently.
Snails collect their metabolic wastes in the coelom -which has been reduced to a sac surrounding the heart. Ammonia, and other wastes, are then funneled down metanephridial tubules where vital salts are reabsorbed. The remainder of waste (urine) is released into the mantle cavity where currents flush it out into the environment.
Reproduction/Development
The prosobranchs and pulmonates have different modes of reproduction. Prosobranchs have separate male and female sexes. The males often have a copulatory organ, either a penis or a tentacle that is modified for the task of delivering sperm. Some females release fertilized eggs to hatch in the environment while others give birth to live young. The females of some species can also reproduce by parthenogenesis. Researchers believe that these gastropods reproduce in this manner in order to colonize habitats where the chance of finding a mate is unlikely. Pulmonates are hermaphroditic with both female and male reproductive parts in one animal. Individuals of some species can self-fertilize, but pay the price of slower maturation and lower fecundity. Other species must find a mate because male and female reproductive organs mature gametes at different points in time.
Unlike their marine counterparts, freshwater snails never have a free-swimming larval stage. Young snails simply look like smaller versions of their parents. To compensate for the absence of this life stage, freshwater snails develop at an accelerated rate and reproduce at a younger age. Pulmonates and prosobranchs have marked developmental differences. Pulmonates are r-strategists. Members are opportunists and colonizers -proven by the fact that they ventured onto land, were able to survive, and then re-adapt to an aquatic environment. By contrast prosobranchs are k-strategists with longer lifespans, low fecundity, and delayed maturation.
Ecology
Most freshwater snails are herbivores or detritevores (organic matter feeders). They employ their radula, a tongue-shaped structure with many tiny tooth-like projections to scrape food from surfaces. It is then retracted back into the mouth and the food is passed from the radula to the esophagus so it can travel to the gut for digestion. If the radula loosens a large chunk of food, then the snail rubs it along a hard plate at the roof of its mouth to break it into smaller pieces. The snail groups also have a special enzyme type, created by the snail version of a liver, called cellulase that is found in very few other invertebrates but plays an important role in dissolving the cell walls of algae and detritus.
The distribution of snails is limited by a variety of different factors. However, the most important chemical factors are pH and the calcium carbonate content of water. Forty five percent of freshwater snails live in water that has a calcium concentration of 20 mg/l or more. Ninety five percent require a concentration of no less than 3 mg/l.
 (Source: Biodiversity Institute of Ontario)
(Source: Biodiversity Institute of Ontario) A pond that is prone to drying up in the summer, or freezing in the winter will have a lower diversity of species than more permanent habitats. The distribution of pulmonates is often described as "passive", which implies that these snails are happy to stay where they are unless a current comes along and sweeps them downstream.
Interspecific competition does not seriously limit snail distributions. There is very rarely any competition between snail species, since many are fairly specialized in their prey choice.
The species that predate snails can be divided into two groups: the shell crushers and the shell invaders. The largest group is the shell crushing predators. Pumpkinseed sunfish and mud minnows use their tough pharyngeal/ teeth to crack open snail shells, while crayfish use their mandibles (chewing mouthparts) to accomplish the same end. Shell-invading predators such as smaller crayfish, leeches, and belastomatids (giant water bugs) are not strong enough to crush shells, but they manage to devour the snail by pulling them out of their shells.
Idiosyncratic inverts
Snails have a wide range of defensive tactics to avoid being captured and eaten. The type that partially dig into sediments simply thicken their shells if the calcium is available. Researchers have found that predators seek out weaker-shelled prey instead of tackling a tougher prey. Speed is an attribute not usually associated with these creatures yet some snails can make a quick escape by crawling above the surface of the water. This involves some fancy "foot"work. Other snails react to attack by shaking their shell so a potential predator can't grab it.
 Image by H. Zell
Image by H. Zell