Food Web (Ecology)
Food Web
| Topics: |
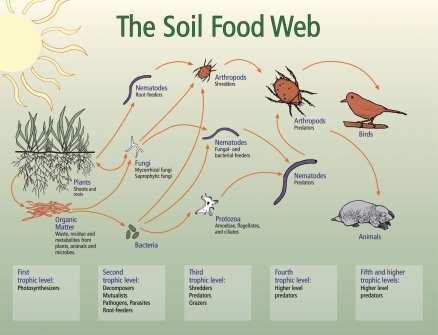
A food web is a graphical description of feeding relationships among species in an ecological community, that is, of who eats whom (Fig. 1). It is also a means of showing how energy and materials (e.g., carbon) flow through a community of species as a result of these feeding relationships. Typically, species are connected by lines or arrows called "links", and the species are sometimes referred to as "nodes" in food web diagrams.
The pioneering animal ecologist Charles Elton (1927) introduced the concept of the food web (which he called food cycle) to general ecological science. As he described it: "The herbivores (Herbivory) are usually preyed upon by carnivores, which get the energy of the sunlight (Solar radiation) at third-hand, and these again may be preyed upon by other carnivores, and so on, until we reach an animal which has no enemies, and which forms, as it were, a terminus on this food cycle. There are, in fact, chains of animals linked together by food, and all dependent in the long run upon plants. We refer to these as 'food-chains', and to all the food chains in a community as the 'food-cycle.'"
A food web differs from a food chain in that the latter shows only a portion of the food web involving a simple, linear series of species (e.g., predator, herbivore, plant) connected by feeding links. A food web aims to depict a more complete picture of the feeding relationships, and can be considered a bundle of many interconnected food chains occurring within the community. All species occupying the same position within a food chain comprise a trophic level within the food web. For instance, all of the plants in the foodweb comprise the first or "primary producer" tropic level, all herbivores (Herbivory) comprise the second or "primary consumer" trophic level, and carnivores that eat herbivores (Herbivory) comprise the third or "secondary consumer" trophic level. Additional levels, in which carnivores eat other carnivores, comprise a tertiary trophic level.
Elton emphasized early on that food chains tend to show characteristic patterns of increasing body size as one moves up the food chain, for example from phytoplankton to invertebrate grazers to fishes, or from insects to rodents to larger carnivores like foxes. Because individuals of small-bodied species require less energy and food than individuals of larger-bodied species, a given amount of energy can support a greater number of individuals of the smaller-bodied species. Hence, ecological communities typically show what Elton called a pyramid of numbers (later dubbed the Eltonian pyramid), in which the species at lower trophic levels in the food web tend to be more numerous than those at higher trophic levels.

A second reason for the pyramid of numbers is low ecological efficiency: some energy is lost at each transfer between consumer and prey, such that the energy that reaches top predators is a very small fraction of that available in the plants at the base of the food web. Although there is wide variation among types of organisms and types of ecosystems, a general rule of thumb is that available energy decreases by about an order of magnitude at each step in the food chain. That is, only about 10% of the energy harvested by plants is consumed and converted into herbivore biomass, only 10% of that makes it into biomass of primary carnivores, and so on. Thus, the structure of food webs is dictated in part by basic constraints set by thermodynamics. The predictable dissipation of energy at each step in food chains is one of the factors thought to limit the length of most food chains to a maximum of four or five steps. Cohen et al. (2003) emphasized that the correlations among body size, abundance, and trophic level produce a characteristic trivariate structure to (pelagic) food webs (Fig. 2).
The pyramid of numbers is less obvious at the most basal levels in terrestrial communities based on trees, which are typically much larger than the herbivores (Herbivory) that feed on them. Pyramids of numbers or biomass may even be inverted in cases where the microscopic plants that support the web show very rapid turnover, that is, where they grow and are eaten so rapidly that there is less plant biomass than herbivore biomass present at a given time.
Decomposers are an assemblage of small organisms, including invertebrates, fungi, and bacteria, that do not fit neatly into any trophic level because they consume dead biomassof organisms from all trophic levels. Decomposers are a critical component of the food web, however, because they recycle nutrients that otherwise would become sequestered in accumulating detritus.
References and further reading
- Cohen, J., T. Jonsson, and S.R. Carpenter. 2003. Ecological community description using the food web, species abundance, and body size. Proceedings of the National Academy of Sciences USA. 100: 1781–1786.
- Elton, C. 1927. Animal ecology. Sidgwick and Jackson Ltd, London, UK.
- Hardy, A.C. 1924. The herring in relation to its animate environment, Part I. Ministry of Agriculture and Fisheries, Fishery Investigations, Series 2, Volume 7, number 3.
- Lindemann, R.L. 1942. The trophic-dynamic aspect of ecology. Ecology 23: 399-418.
- May, R.M. 1973. Stability and complexity in model food webs. Princeton University Press, Princeton, NJ.
- Morin, P.J. 1999. Community ecology. Blackwell Science, Oxford, UK.
- Paine, R.T. 1980. Food Webs: Linkage, Interaction Strength and Community Infrastructure. Journal of Animal Ecology 49:666-685.
- Pimm, S.L. 1982. Food webs. Chapman and Hall, London.